| Психология | Соматология | Математика | Физика | Химия | Наука Общая лексика |
| А | Б | В | Г | Д | Е | Ж | З | И | К | Л | М | Н | О | П | Р | С | Т | У | Ф | Х | Ц | Ч | Ш | Щ | Э | Ю | Я |
| A | B | C | D | E | F | G | H | I | J | K | L | M | N | O | P | Q | R | S | T | U | V | W | X | Y | Z |
|
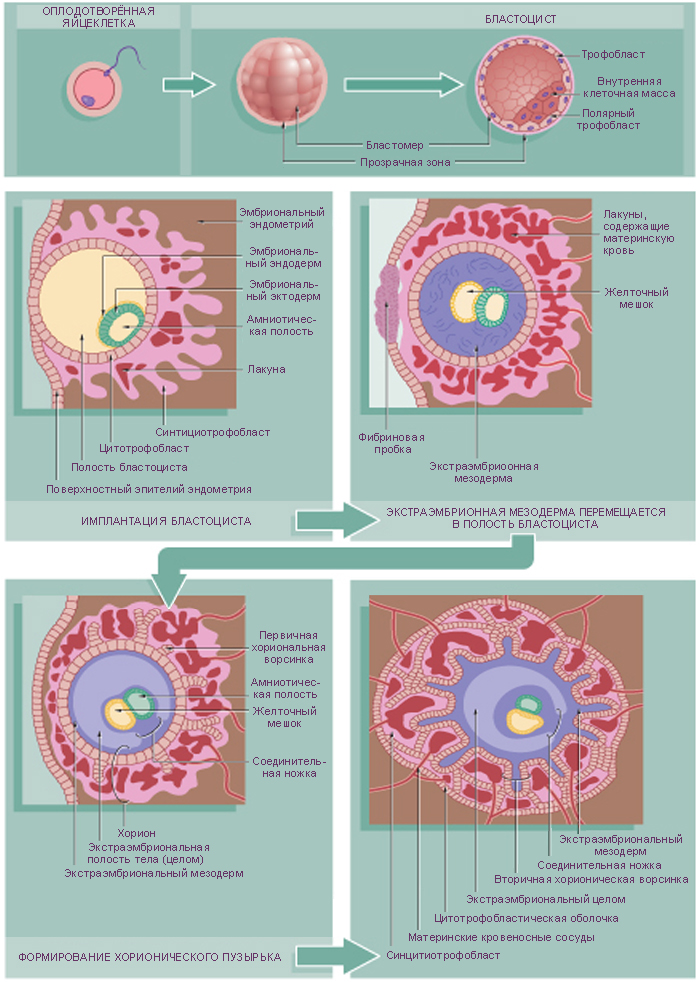
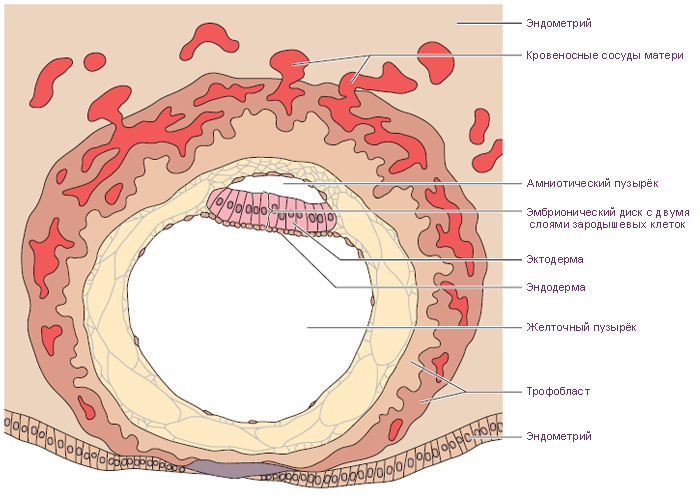
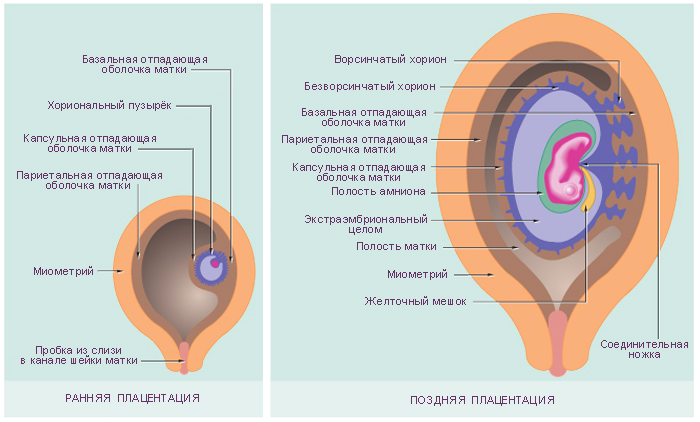
Внезародышевые органы, развивающиеся в процессе эмбриогенеза вне
тела зародыша, выполняют многообразные функции, обеспечивающие рост
и развитие самого зародыша. Некоторые из этих органов, окружающих
зародыш, называют также зародышевыми оболочками. К этим органам
относятся амнион, желточный мешок, аллантоис, хорион, плацента (рис. 45).
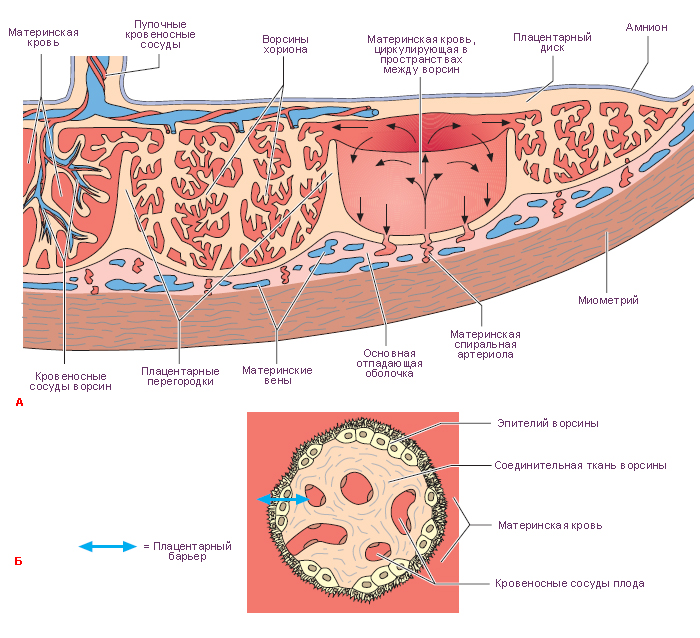
Плацента
Плацента (детское место) человека относится к типу дискои-
дальных гемохориальных ворсинчатых плацент (см. рис. 47; рис. 48). Это
важный временный орган с многообразными функциями (см. с. 101),
которые обеспечивают связь плода с материнским организмом. Вместе с тем
плацента создает барьер между кровью матери и плода. Плацента состоит
из двух частей: зародышевой, или плодной (pars fetalis), и материнской (pars
materna). Плодная часть представлена ветвистым хорионом и приросшей
к нему изнутри амниотической оболочкой, а материнская —
видоизмененной слизистой оболочкой матки, отторгающейся при родах (decidua
basalis).
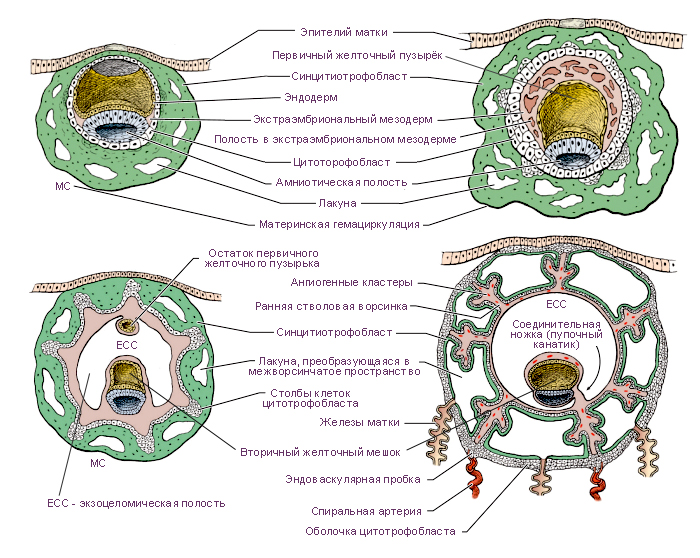
Развитие плаценты начинается на 3-й неделе, когда во вторичные
ворсины начинают врастать сосуды и образовываться третичные ворсины, и
заканчивается к концу 3-го месяца беременности. На 6—8-й неделе вокруг
сосудов дифференцируются элементы соединительной ткани. В дифферен-
цировке фибробластов и синтезе ими коллагена важную роль играют
витамины А и С, без достаточного поступления которых в организм
беременной женщины нарушается прочность связи зародыша с материнским
организмом и создается угроза самопроизвольного аборта.
В основном веществе соединительной ткани хориона содержится
значительное количество гиалуроновой и хондроитинсерной кислот, с
которыми связана регуляция проницаемости плаценты.
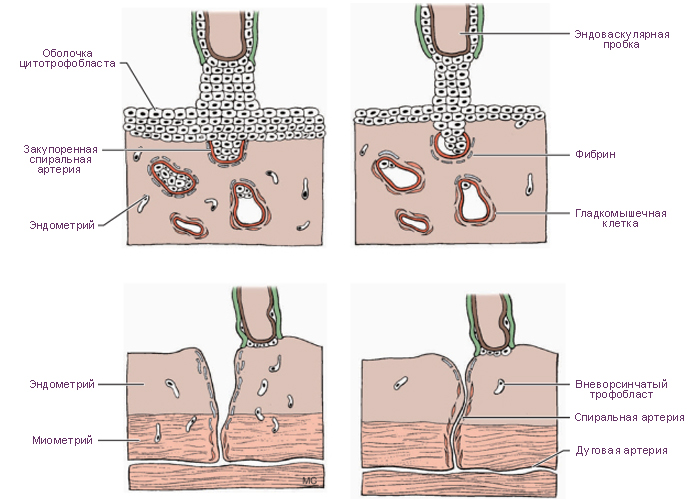
При развитии плаценты происходят разрушение слизистой оболочки
матки, обусловленное протеолитической активностью хориона, и смена
гистиотрофного питания на гематотрофное. Это означает, что ворсины
хориона омываются кровью матери, излившейся из разрушенных сосудов
эндометрия в лакуны. Однако кровь матери и плода в нормальных условиях
никогда не смешивается.
Гематохориальный барьер, разделяющий оба кровотока,
состоит из эндотелия сосудов плода, окружающей сосуды соединительной
ткани, эпителия хориальных ворсин (цитотрофобласт и симпластотрофоб-
ласт), а кроме того, из фибриноида, который местами покрывает ворсины
снаружи.
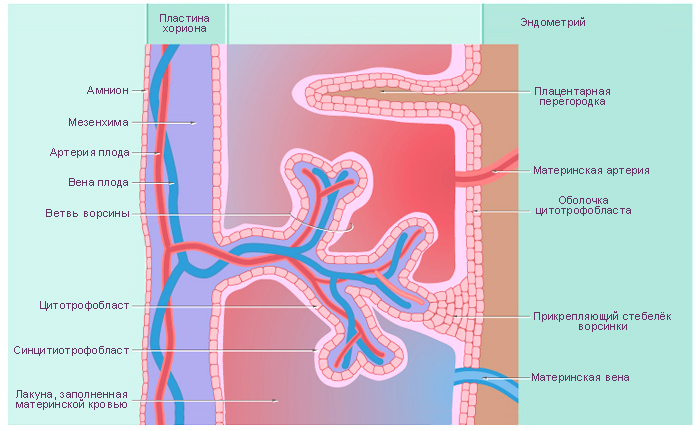
Зародышевая, или плодная, часть плаценты к концу 3-го
месяца представлена ветвящейся хориальной пластинкой, состоящей из
волокнистой (коллагеновой) соединительной ткани, покрытой цито- и симпла-
стотрофобластом (многоядерная структура, покрывающая редуцирующийся
цитотрофобласт). Ветвящиеся ворсины хориона (стволовые, якорные)
хорошо развиты лишь со стороны, обращенной к миометрию. Здесь они про-
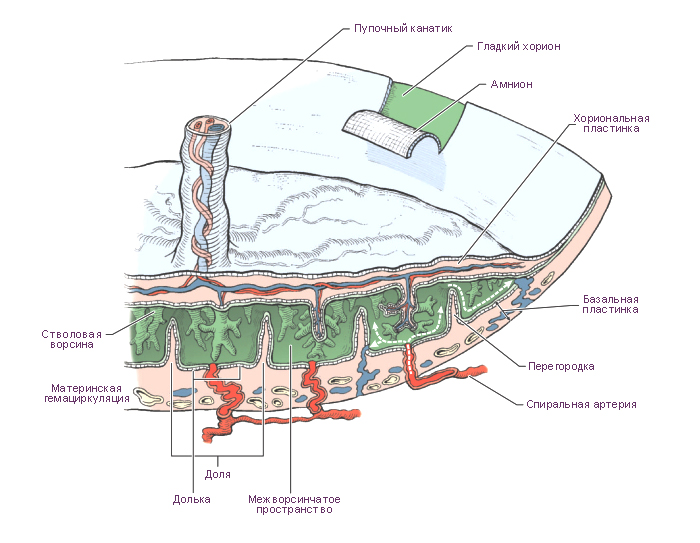
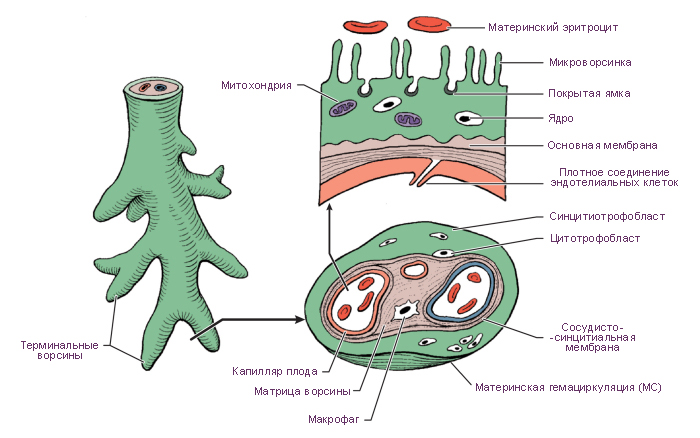
Рис. 48. Плацента гемохориального типа. Динамика развития ворсин хориона.
А — строение плаценты (стрелками указана циркуляция крови в сосудах и в одной из лакун,
где удалена ворсинка): 1 — эпителий амниона; 2 — хориальная пластинка; 3 — ворсинка;
4 — фибриноид; 5 — желточный мешок; 6 — пупочный канатик; 7 — перегородка плаценты;
8 — лакуна; 9 — спиральная артерия; 10 — базальный слой эндометрия; 11 — миометрий;
Б — строение первичной ворсины трофобласта A-я неделя);
В — строение вторичной эпителиально-мезенхимальной ворсины хориона B-я неделя);
Г _ строение третичной ворсины хориона — эпителиально-мезенхимальной с кровеносными
сосудами C-я неделя);
Д — строение ворсины хориона C-й месяц);
Е — строение ворсин хориона (9-й месяц): 1 — межворсинчатое пространство; 2 —
микроворсинки; 3 — симпластотрофобласт; 4 — ядра симпластотрофобласта; 5 — цитотрофобласт;
6 — ядро цитотрофобласта; 7 — базальная мембрана; 8 — межклеточное пространство; 9 —
фибробласт; 10 — макрофаги (клетки Кащенко—Гофбауэра); 11 — эндотелиоцит; 12 —
просвет кровеносного сосуда; 13 — эритроцит; 14 — базальная мембрана гемокапилляра (по
Э. М. Швирсту).
Рис. 49. Срез ворсины хориона
17-суточного зародыша человека
(«Крым»). Микрофотография.
I — симпластотрофобласт; 2 — цито-
трофобласт; 3 — мезенхима хориона
(по Н.Р.Барсукову).
ходят через всю толщу плаценты и
своими вершинами погружаются в
базальную часть разрушенного
эндометрия.
Хориальный эпителий, или
цитотрофобласт, на ранних
стадиях развития представлен
однослойным эпителием с овальными
ядрами. Эти клетки размножаются ми-
тотическим путем. Из них
развивается симпластотрофобласт. В сим-
пластотрофобласте содержится
большое количество различных
протеолитических и окислительных ферментов (АТФазы, щелочная и
кислая фосфатазы, 5-нуклеотидазы, ДПН-диафоразы, глюкозо-6-фосфатдегид-
рогеназы, а-ГФДГ, сукцинатдегидрогеназа — СДГ, цитохромоксидаза —
ЦО, моноаминоксидаза — МАО, неспецифические эстеразы, ЛДГ, НАД-
и НАДФ-диафоразы и др. — всего около 60), что связано с его ролью в
обменных процессах между организмом матери и плода. В цитотрофобласте
и в симпласте выявляются пиноцитозные пузырьки, лизосомы и другие
органеллы. Начиная со 2-го месяца, хориальный эпителий истончается и
постепенно заменяется симпластотрофобластом. В этот период
симпластотрофобласт по толщине превосходит цитотрофобласт, на 9—10-й неделе
симпласт истончается, а количество ядер в нем увеличивается. На
поверхности симпласта, обращенной в лакуны, появляются многочисленные
микроворсинки в виде щеточной каемки (см. рис. 48; рис. 49, 50).
Между симпластотрофобластом и клеточным трофобластом имеются
щелевидные субмикроскопические пространства, доходящие местами до
базальной мембраны трофобласта, что создает условия для двустороннего
проникновения трофических веществ, гормонов и др.
Во второй половине беременности и особенно в конце ее трофобласт
сильно истончается и ворсины покрываются фибриноподобной оксифиль-
ной массой, являющейся продуктом свертывания плазмы и распада
трофобласта («фибриноид Лангханса»).
С увеличением срока беременности уменьшается количество
макрофагов и коллаген продуцирующих дифференцированных фибробластов,
появляются фиброциты. Количество коллагеновых волокон, хотя и нарастает, но
до конца беременности в большинстве ворсин остается незначительным.
Большая часть стромальных клеток (миофибробластов) характеризуется уве-
Рис. 50. Плацентарный барьер на 28-й неделе беременности. Электронная
микрофотография, х 45 000 (по У.Ю.Яцожинской).
1 — симпластотрофобласт; 2 — цитатрофобласт; 3 — базальная мембрана трофобласта, 4 —
базапьная мембрана эндотелия; 5 — эндотелиоцит; 6 — эритроцит в капилляре.
личенным содержанием цитоскелетных сократительных белков (виментин,
десмин, актин и миозин).
Структурно-функциональной единицей сформированной плаценты
является котиледон, образованный стволовой («якорной») ворсиной и ее
вторичными и третичными (конечными) разветвлениями. Общее количество
котиледонов в плаценте достигает 200.
Материнская часть плаценты представлена базальной пластинкой
и соединительнотканными септами, отделяющими котиледоны друг от
друга, а также лакунами, заполненными материнской кровью. В местах
контакта стволовых ворсин с отпадающей оболочкой встречаются также трофо-
бластические клетки (периферический трофобласт).
Уже на ранних стадиях беременности ворсины хориона разрушают
ближайшие к плоду слои основной отпадающей оболочки, и на их месте обра-
зуются заполненные материнской кровью лакуны, в которые свободно
свисают ворсины хориона.
Глубокие неразрушенные части отпадающей оболочки вместе с трофо-
бластом образуют базальную пластинку.
Базальный слой эндометрия (lamina basalis) — соединительная ткань
слизистой оболочки матки, содержащая децидуалъные клетки. Эти крупные,
богатые гликогеном клетки соединительной ткани расположены в глубоких
слоях слизистой оболочки матки. Они имеют четкие границы, округлые ядра
и оксифильную цитоплазму. В течение 2-го месяца беременности децидуаль-
ные клетки значительно укрупняются. В их цитоплазме, кроме гликогена,
выявляются липиды, глюкоза, витамин С, железо, неспецифические эсте-
разы, дегидрогеназа янтарной и молочной кислот. В базальной пластинке,
чаще в месте прикрепления ворсин к материнской части плаценты,
встречаются скопления клеток периферического цитотрофобласта. Они
напоминают децидуальные клетки, но отличаются более интенсивной базофилией
цитоплазмы. Аморфная субстанция (фибриноид Рора) находится на
поверхности базальной пластинки, обращенной к хориальным ворсинам.
Фибриноид играет существенную роль в обеспечении иммунологического гомео-
стаза в системе мать — плод.
Часть основной отпадающей оболочки, расположенной на границе
ветвистого и гладкого хориона, т.е. по краю плацентарного диска, при
развитии плаценты не разрушается. Плотно прирастая к хориону, она образует
замыкающую пластинку, препятствующую истечению крови из лакун
плаценты.
Кровь в лакунах непрерывно циркулирует. Она поступает из маточных
артерий, входящих сюда из мышечной оболочки матки. Эти артерии идут
по плацентарным перегородкам и открываются в лакуны. Материнская кровь
оттекает от плаценты по венам, берущим начало от лакун крупными
отверстиями.
Формирование плаценты заканчивается в конце 3-го месяца
беременности. Плацента обеспечивает питание, тканевое дыхание, рост, регуляцию
образовавшихся к этому времени зачатков органов плода, а также его
защиту.
Функции плаценты. Основные функции плаценты: 1) дыхательная,
2) транспорт питательных веществ, воды, электролитов и
иммуноглобулинов, 3) выделительная, 4) эндокринная, 5) участие в регуляции
сокращения миометрия.
Дыхание плода обеспечивается за счет кислорода, присоединенного к
гемоглобину материнской крови, который путем диффузии поступает через
плаценту в кровь плода, где он соединяется с фетальным гемоглобином
(HbF). Связанная с фетальным гемоглобином С02 в крови плода также
диффундирует через плаценту, поступает в кровь матери, где соединяется с
материнским гемоглобином.
Транспорт всех питательных веществ, необходимых для развития плода
(глюкоза, аминокислоты, жирные кислоты, нуклеотиды, витамины,
минеральные вещества), происходит из крови матери через плаценту в кровь
плода, и, наоборот, из крови плода в кровь матери поступают продукты
обмена веществ, выводимые из его организма (выделительная функция).
Электролиты и вода проходят через плаценту путем диффузии и с
помощью пиноцитоза.
В транспорте иммуноглобулинов (Ig) участвуют пиноцитозные
везикулы симпластотрофобласта. Поступивший в кровь плода Ig пассивно
иммунизирует его от возможного действия бактериальных антигенов, которые
могут поступать при заболеваниях матери. После рождения материнский Ig
разрушается и заменяется вновь синтезируемым Ig в организме ребенка при
действии на него бактериальных антигенов. Через плаценту в околоплодные
воды проникают Ig класса G и A (IgG, IgA).
Эндокринная функция является одной из важных, так как плацента
обладает способностью синтезировать и секретировать ряд гормонов,
обеспечивающих взаимодействие зародыша и материнского организма на
протяжении всей беременности. Местом продукции плацентарных гормонов
являются цитотрофобласт и особенно симпластотрофобласт, а также деци-
дуальные клетки.
Одним из первых плацента синтезирует хорионический гонадо-
тропин, концентрация которого быстро нарастает на 2—3-й неделе
беременности, достигая максимума на 8—10-й неделе, причем в крови плода
она в 10—20 раз выше, чем в крови матери. Гормон стимулирует
образование адренокортикотропного гормона (АКТГ) гипофиза, усиливает
секрецию кортикостероидов.
Большое значение в развитии беременности имеет плацентарный
лактоген, который обладает активностью пролактина и лютеотропного
гормона гипофиза. Он поддерживает стероидогенез в желтом теле яичника
в первые 3 мес беременности, а также принимает участие в метаболизме
углеводов и белков. Концентрация его в крови матери прогрессивно
нарастает на 3—4-м месяце беременности и в дальнейшем продолжает
увеличиваться, достигая максимума к 9-му месяцу. Этот гормон совместно с про-
лактином гипофиза матери и плода играет определенную роль в продукции
легочного сурфактанта и фетоплацентарной осморегуляции. Высокая
концентрация его обнаруживается в околоплодных водах (в 10—100 раз
больше, чем в крови матери).
В хорионе, а также в децидуальной оболочке синтезируются
прогестерон и прегнандиол.
Прогестерон (вырабатываемый сначала желтым телом в яичнике, а с
5—6-й недели в плаценте) подавляет сокращения матки, стимулирует ее
рост, оказывает иммунодепрессивное действие, подавляя реакцию
отторжения плода. Около 3/4 прогестерона в организме матери метаболизируется и
трансформируется в эстрогены, а часть выделяется с мочой.
Эстрогены (эстрадиол, эстрон, эстриол) вырабатываются в симпла-
стотрофобласте ворсин плаценты (хориона) в середине беременности, а к
концу беременности их активность усиливается в 10 раз. Они вызывают
гиперплазию и гипертрофию матки.
Кроме того, в плаценте синтезируются меланоцитостимулирующий и
адренокортикотропный гормоны, соматостатин и др.
В плаценте содержатся полиамины (спермин, спермидин), влияющие
на усиление синтеза РНК в гладких мышечных клетках миометрия, а также
на разрушающие их оксидазы. Важную роль играют аминооксидазы (гиста-
миназа, моноаминоксидаза), разрушающие биогенные амины — гистамин,
серотонин, тирамин. Во время беременности их активность возрастает, что
способствует разрушению биогенных аминов и падению концентрации
последних в плаценте, миометрии и крови матери.
Во время родов гистамин и серотонин являются наряду с катехоламинами (нор-
адреналин, адреналин) стимуляторами сократительной деятельности гладких
мышечных клеток (ГМК) матки, и к концу беременности их концентрация
значительно возрастает в связи с резким падением (в 2 раза) активности аминооксидаз (ги-
стаминаза и др.).
При слабой родовой деятельности отмечается усиление активности
аминооксидаз, например гистаминазы (в 5 раз).
Нормальная плацента не является абсолютным барьером для белков. В
частности, а-фетопротеин в конце 3-го месяца беременности проникает в небольшом
количестве (около 10 %) из плода в кровь матери, но на этот антиген материнский
организм не отвечает отторжением, так как во время беременности уменьшается
цитотоксичность материнских лимфоцитов.
Плацента препятствует прохождению ряда материнских клеток и цитотоксичес-
ких антител к плоду. Главную роль в этом играет фибриноид, покрывающий
трофобласт при его частичном повреждении. Это предотвращает поступление в
межворсинчатое пространство плацентарных и плодовых антигенов, а также ослабляет
гуморальную и клеточную «атаку» матери против зародыша.
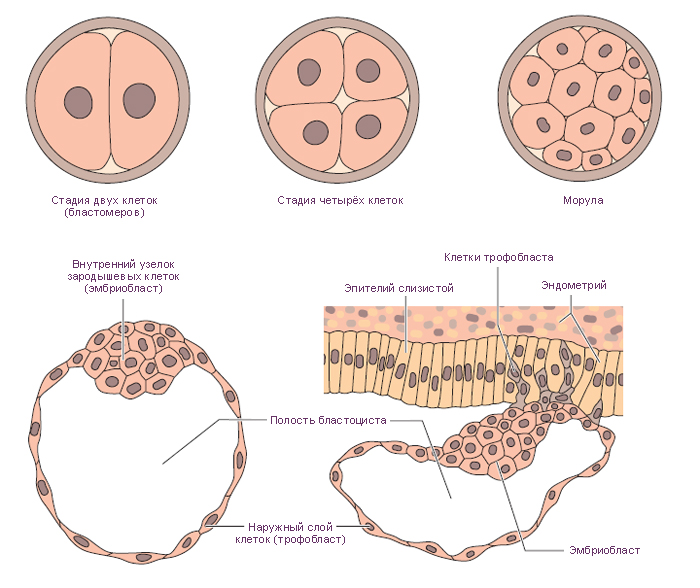
В заключение отметим основные особенности ранних стадий развития
зародыша человека: 1) асинхронный тип полного дробления и образование
«светлых» и «темных» бластомеров; 2) раннее обособление и
формирование внезародышевых органов; 3) раннее образование амниотического
пузырька и отсутствие амниотических складок; 4) наличие в стадии гаструля-
ции двух механизмов — деламинации и иммиграции, в течение которых
происходит также развитие провизорных органов; 5) интерстициальный тип
имплантации; 6) сильное развитие амниона, хориона, плаценты и слабое
развитие желточного мешка и аллантоиса.
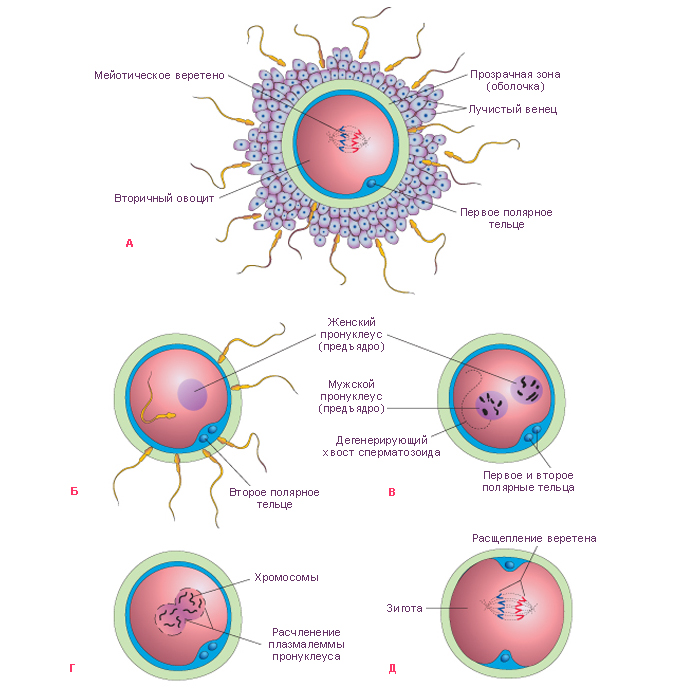
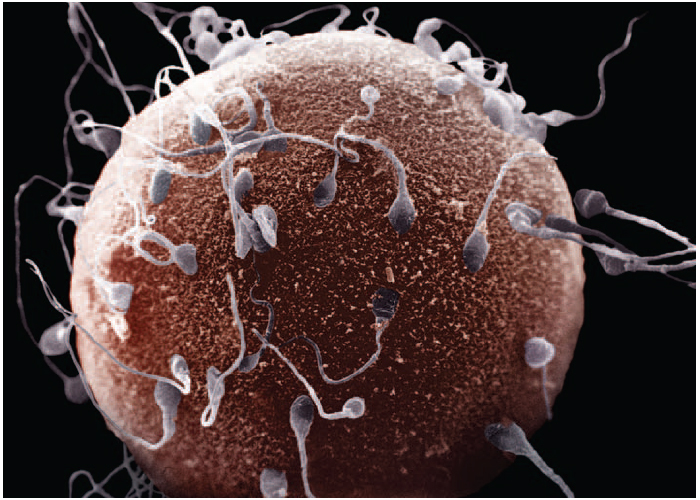
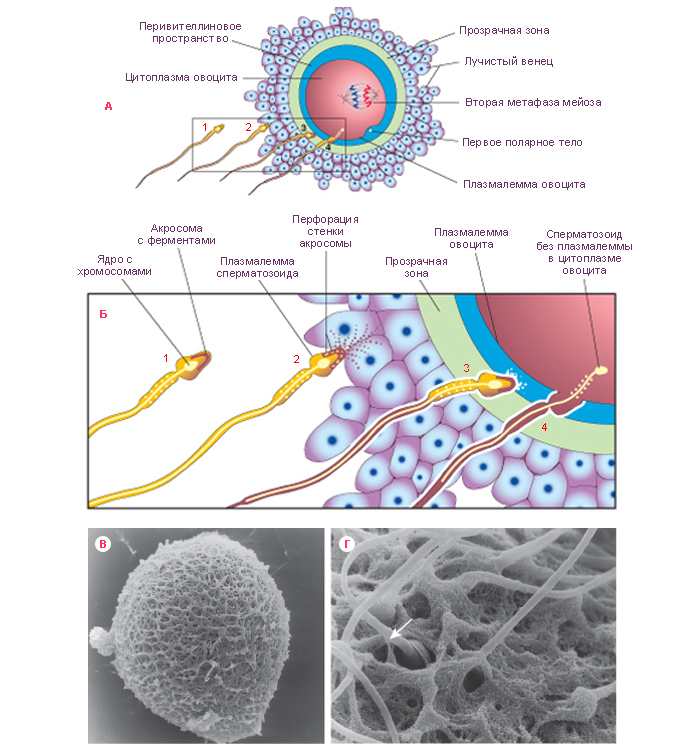
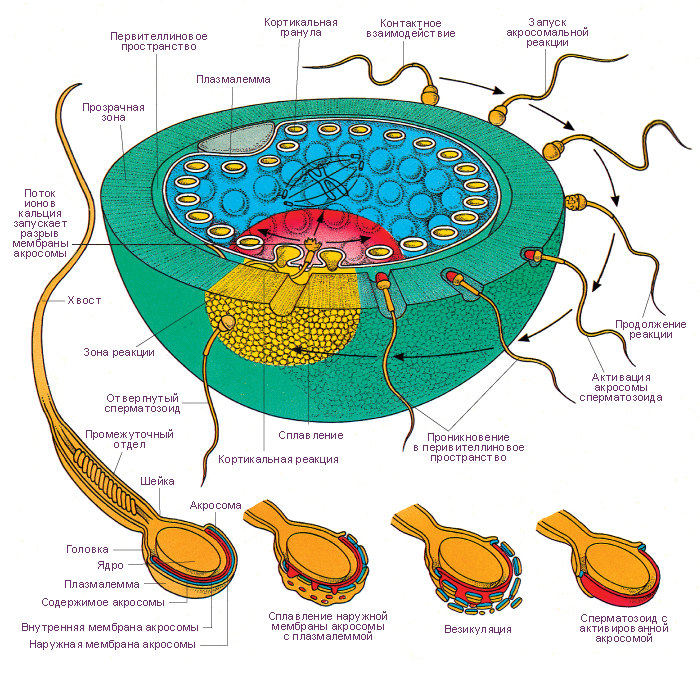
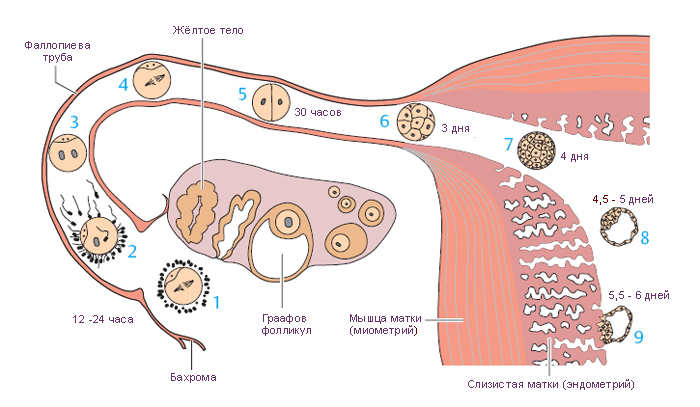
!!!!!!Sexual Selection and the Origins of Human Mating Systems (Oxford Biology) By Alan F. Dixson Publisher: Oxford University Press, USA Number Of Pages: 232 Publication Date: 2009-07-15 2_116/Sexual_Selection_and_Origins_Human_Mating_Systems2009.pdf = рисунок sexact1_1.tif Учебное пособие. . Доступ к данному источнику = Access to the reference. URL: http://www.tryphonov.ru/tryphonov/serv_r.htm#0 quotation Афанасьев с 100 Оплодотворение и образование зиготы Оплодотворение (fertilisatio) — слияние мужской и женской половых клеток, в результате чего восстанавливается диплоидный набор хромосом, характерный для данного вида животных, и возникает качественно новая клетка — зигота (оплодотворенная яйцеклетка, или одноклеточный зародыш). У человека объём эякулята — извергнутой спермы — в норме составляет около 3 мл. Для обеспечения оплодотворения общее количество сперматозоидов в сперме должно быть не менее 150 млн, а концентрация их в 1 мл — 20—200 млн1, хотя в яйцеклетку проникает только один из них, а остальные подготавливают условия для оплодотворения. В половых путях женщины после копуляции их число уменьшается по направлению от влагалища к дистальному концу маточной трубы. В процессе оплодотворения различают три фазы: 1) дистантное взаимодействие и сближение гамет; 2) контактное взаимодействие и активизация яйцеклетки; 3) вхождение сперматозоида в яйцо и последующее слияние — сингамия. Первая фаза — дистантное взаимодействие — обеспечивается хемотаксисом — совокупностью специфических факторов, повышающих вероятность столкновения половых клеток. Важную роль в этом играют гамоны — химические вещества, вырабатываемые половыми клетками (рис. 32). Установлено, что яйцеклетки выделяют пептиды, способствующие привлечению сперматозоидов. Сразу после эякуляции спермин неспособны к проникновению в яйцеклетки до тех пор, пока не произойдет капацитация — приобретение спермиями оплодотворяющей способности под действием секрета женских половых путей, которое длится 7 ч. В процессе капацитации с плазмолеммы спермия в области акросомы удаляются гликопротеины и протеины семенной плазмы, что способствует акросомальной реакции. В механизме капацитации большое значение принадлежит гормональным факторам, прежде всего прогестерону (гормон желтого тела), активизирующему секрецию железистых клеток яйцеводов. Во время капацитации происходят связывание холестерина цитолеммы спермия альбуминами женских половых путей и обнажение рецепторов половых клеток. Оплодотворение происходит в ампулярной части яйцевода. Оплодотворению предшествует осеменение — взаимодействие и сближение гамет (дистантное взаимодействие), обусловленное хемотаксисом. Вторая фаза оплодотворения — контактное взаимодействие, во время которого сперматозоиды вращают яйцеклетку. Многочисленные спермин приближаются к яйцеклетке и вступают в контакт с ее оболочкой. Яйцеклетка начинает совершать вращательные движения вокруг своей оси со скоростью 4 вращения в минуту. Эти движения обусловлены влиянием биения жгутиков сперматозоидов и продолжаются около 12 ч. В процессе взаимодействия мужской и женской половых клеток в спермиях происходит акросомальная р е а к ц и я. Она заключается в слиянии наружной мембраны акросомы с передними 2/3 плазмолеммы спермия. Затем Рис. 32. Дистантное и контактное взаимодействие спермиев и яйцеклетки. 1 — сперматозоид и его рецепторы на головке; 2 — отделение углеводов с поверхности го- ловки при капацитацин; 3 — связывание рецепторов сперматозоида с NAP-рецепта рам и яйцеклетки (N - ацети л - гл ю коза мин- рецептор, заключенный в Zp3); 4 — Zp3 (третья фракция блестящей зоны)! 5 — цитолемма яйцеклетки; ГП, ГГП — гиногамоны; АП, АГЛ — андро- гамоны. в области слияния мембраны разрываются и ферменты акросомы выходят в окружающую среду. Инициация второй фазы оплодотворения происходит под влиянием сульфати- рованных полисахаридов блестящей зоны, которые вызывают поступление ионов кальция и натрия в головку спермия, замещение ими ионов калия и водорода и разрыв мембраны акросомы. Прикрепление спермия к яйцеклетке происходит под влиянием углеводной группы фракции гл и коп роте и нов прозрачной зоны яйцеклетки. Рецепторы спермия для прозрачной зоны представляют собой фермент глико- зилтрансферазу, находящийся на поверхности акросомы головки, который «узнает* сахар N-ацети л глюкозам и н — рецептор женской половой клетки. Плазматические мембраны в месте контакта половых клеток сливаются и происходит плазмогамия — объединение цитоплазм обеих гамет. Сперматозоиды при контакте с яйцеклеткой могут связывать десятки тысяч молекул гликопротеида Zp3. При этом отмечается запуск акросомаль- ной реакции. Акросомальная реакция характеризуется повышением проницаемости плазмолеммы спермия к ионам Сан, деполяризацией ее, что способствует слиянию плазмолеммы с передней мембраной акросомы. Блестящая зона оказывается в непосредственном контакте с акросомальными ферментами. Ферменты разрушают блестящую зону, спермий проходит через нее и входит в перивителлиновое пространство, расположенное между блестящей зоной и плазмолеммой яйцеклетки. Через несколько секунд изменяются свойства плазмолеммы яйцеклетки и начинается кортикальная реакция, а через несколько минут изменяются свойства блестящей зоны — Zp (зонная реакция). Рис. 33. Оплодотворение (по Вассерману с изменениями). 1, 2, 3, 4 — стадии акросомной реакции; 5 — zonn pellucida (блестящая зона); 6 — перивител- л и новое пространство: 7 — плазматическая мембрана; 8 — кортикальная гранула; 8а -¦ кортикальная реакция; 9 — вхождение спермия в яйцеклетку; 10 — зонная реакция. У млекопитающих при оплодотворении в яйцеклетку проникает лишь один сперматозоид. Такое явление называется моноспермие й. Оплодотворению способствуют сотни других принимающих участие в осеменении сперматозоидов. Ферменты, выделяемые из акросом, — спермолизины (трипсин, гиалуронидаза) разрушают лучистый венец, расщепляют глико- заминогликаны прозрачной зоны яйцеклетки. Отделяющиеся фолликулярные клетки склеиваются в конгломерат, который вслед за яйцеклеткой перемещается по трубе благодаря мерцанию ресничек эпителиальных клеток слизистой оболочки. Третья фаза. В ооплазму проникают головка и промежуточная часть хвостового отдела. После вхождения сперматозоида в яйцеклетку на периферии ооплазмы происходит уплотнение ее (зонная реакция) и образуется оболочка оплодотворения. Кортикальная реакция — слияние плазмолеммы яйцеклетки с мембранами кортикальных гранул, в результате чего содержимое из гранул выходит в пери вителл и новое пространство и воздействует на молекулы гли- копротеидов блестящей зоны (рис. 33). Вследствие этой зонной реакции мо- Рнс. 34. Фазы оплодотворения и начало дробления (схема). 1 — ооплазма; la — кортикальные гранулы; 2 — ядро; 3 — блестящая зона; 4 — фолликулярный эпителий; 5 — спермин; 6 — редукционные тельца; 7 — митотическое деление ооиита; 8 — бугорок оплодотворения; 9 — оболочка оплодотворения; 10 — женский пронуклеус; 11 — мужской пронуклеус; 12 — синкарион; 13 — первое митотическое деление зиготы; 14 — бла- стомеры. лекулы Zp3 модифицируются и утрачивают способность быть рецепторами спермиев. Образуется оболочка оплодотворения толщиной 50 нм, препятствующая полиспермии — проникновению других спермиев. Механизм кортикальной реакции включает приток ионов натрия через участок мембраны сперматозоида, встроенный в поверхность яйцеклетки после завершения акросомальной реакции. В результате отрицательный мембранный потенциал клетки становится слабоположительным. Приток ионов натрия обусловливает высвобождение ионов кальция из внутриклеточных депо и увеличение его содержания в гиалоплазме яйцеклетки. Вслед за этим начинается экзоцитоз кортикальных гранул. Высвобождающиеся из них про- теолитические ферменты разрывают связи между блестящей зоной и плаз- молеммой яйцеклетки, а также между спермиями и прозрачной зоной. Кроме того, выделяется гликопротеид, связывающий воду и привлекающий ее в пространство между плазмолеммой и блестящей зоной. Вследствие этого формируется перивитеминоеое пространство. Наконец, выделяется фактор, способствующий затвердению прозрачной зоны и образованию из нее оболочки оплодотворения. Рис. 35. Яйцеклетка и зигота человека (по Б.П.Хватову). А — яйцеклетка человека после овуляции: 1 — цитоплазма; 2 — ядро; 3 — блестящая зона; 4 — фолликулярные клетки, образующие лучистый венец. Б — зигоТа человека в стадии сближения мужского и женского ядер (пронуклеусов): I — женское ядро; 2 — мужское ядро. Благодаря механизмам предотвращения полиспермии только одно гаплоидное ядро сперматозоида получает возможность слиться с одним гаплоидным ядром яйцеклетки, что приводит к восстановлению характерного для всех клеток диплоидного набора. Проникновение сперматозоида в яйцеклетку через несколько минут значительно усиливает процессы внутриклеточного обмена, что связано с активизацией ферментативных ее систем. Взаимодействие сперматозоидов с яйцеклеткой может быть заблокировано при помощи антител против веществ, входящих в прозрачную зону. На этом основании изыскиваются способы иммунологической контрацепции. После сближения женского и мужского пронуклеусов, которое продолжается у млекопитающих около 12 ч, образуется зигота — одноклеточный зародыш (рис. 34, 35, А, Б). Уже на стадии зиготы выявляются презумптив- ные зоны (лат. presumptio — вероятность, предположение) как источники развития соответствующих участков бластулы, из которых в дальнейшем формируются зародышевые листки1. Дробление и образование бластулы Дробление (fissio) — последовательное митотическое деление зиготы на клетки (бластомеры) без роста дочерних клеток до размеров материнской. Образующиеся бластомеры остаются объединенными в единый организм зародыша. В зиготе образуется митотическое веретено между отдаляющимися к полюсам центриолями, внесенными сперматозоидом. Пронуклеусы вступают в стадию профазы с формированием объединенного диплоидного набора Метод выявления презумптивных зон предложен немецким эмбриологом Фогтом. Рис. 36. Зародыш человека на ранних стадиях развития (по Гертигу и Рокку). А — стадия двух бластомеров; Б — бластоциста: [ — эмбриобласт; 2 — чрофобласт; 3 — полость бласто цисты. хромосом яйцеклетки и сперматозоида. Пройдя все остальные фазы митоти- ческого деления, зигота разделяется на две дочерние клетки — бластомеры (от греч. blastos — зачаток, meros — часть). Вследствие фактического отсутствия G,-периода, во время которого происходит рост клеток, образовавшихся в результате деления, клетки гораздо меньше материнской, поэтому и величина зародыша в целом в этот период независимо от числа составляющих его клеток не превышает величину исходной клетки — зиготы. Все это позволило назвать описываемый процесс дроблением (т. е. измельчением), а клетки, образующиеся в процессе дробления, — бластомерами. Дробление зиготы человека начинается к концу первых суток и характеризуется как полное неравномерное асинхронное. В течение первых суток оно происходит медленно. Первое дробление (деление) зиготы завершается через 30 ч, б результате образуется 2 бластомера, покрытых оболочкой оплодотворения. За стадией двух бластомеров следует стадия трех бластомеров. С первых же дроблений зиготы формируются два вида бластомеров — «темные» и «светлые». «Светлые», более мелкие, бластомеры дробятся быстрее и располагаются одним слоем вокруг крупных «темных», которые оказываются в середине зародыша. Из поверхностных «светлых» бластомеров в дальнейшем возникает трофобласт, связывающий иродыш с материнским организмом и обеспечивающий его питание. Внутренние, «темные», бластомеры формируют эмбриобласт, из которого образуются тело зародыша и некоторые внеэародышевые органы (амнион, желточный мешок, аллан- тоис). Начиная с трех суток, дробление идет быстрее, и на 4-е сутки зародыш состоит из 7—12 бластомеров. Уже через 50—60 ч образуется плотное скопление клеток — морула, а на 3—4-е сутки начинается формирование бластоцисты — полого пузырька, заполненного жидкостью (рис. 36, 37). Бластоциста в течение 3 сут перемещается по яйцеводу к матке и через 4 сут попадает в матку. Бластоциста находится в полости матки в свободном 105 Рис. 37. Дробление, гаструляция и имплантация зародыша человека (схема). I — дробление; 2 — морула; 3 — бластоциста; 4 — полость бластоцисты; 5 — эмбриобласт; 6 — трофобласт; 7 — зародышевый узелок: а — эпибласт, б — гипобласт; 8 — оболочка оплодотворения; 9 — имниотический (эктодермольный) пузырек; 10 — внеэародышевая мезодерма; II — эктодерма; 12 — энтодерма; 13 — цитотрофобласт; 14 — симпластотрофобласт; 15 — зародышевый диск; 16 — лакуны с материнской кровью; 17 — хорион; 18 — амниоти- ческая ножка; 19 — желточный пузырек; 20 — слизистая оболочка матки; 21 — яйцевод. виде в течение 2 дней E-е и 6-е сутки), и эта стадия обозначается как свободная бластоциста. К этому времени бластоциста увеличивается благодаря росту числа бластомеров — клеток эмбриобласта и трофобласта — до 100 и более вследствие усиленного всасывания трофобластом секрета маточных желез, а также вследствие активной выработки жидкости самим трофобластом (см. рис. 37). Эмбриэбласт располагается в виде узелка зародышевых клеток («зародышевый узелок*), который прикрепляется изнутри к трофобласту на одном из полюсов бластоцисты и начинается имплантация. Имплантация Имплантация (лат. implantatio — врастание, укоренение) — внедрение зародыша в слизистую оболочку матки. Различают две стадии имплантации: адгезию (прилипание), когда зародыш прикрепляется к внутренней поверхности матки, и инвазию 106
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Санкт-Петербург, Россия, 1996-2015
Разрешается некоммерческое цитирование материалов данной энциклопедии при условии
полного указания источника заимствования: имени автора, названия и WEB-адреcа данной энциклопедии
| Автор | Новые идеи | Индекс | Библиотека | Поиск | Отзывы | Ссылка | Пожертвования | Параллельный сайт |
Всего посетителей = Altogether Visitors :
Посетителей раздела «Соматология» = Visitors of section «Somatlogy» : |