Таблица. Некоторые органические анионы, секретируемые в жидкость проксимальных мочевых канальцев.
Таблица. Некоторые органические катионы, секретируемые в жидкость проксимальных мочевых канальцев.
12333. Каналъцевая секреция
В выделении продуктов обмена и чужеродных веществ имеет значение их секреция из крови в просвет канальца против концентрационного и электрохимического градиентов. Этот дополнительный механизм выделения ряда веществ, помимо их фильтрации в клубочках, позволяет быстро экскретировать некоторые органические кислоты и основания, а также некоторые ионы, например К+. Секреция органических кислот (феноловый красный, ПАГ, диодраст, пенициллин) и органических оснований (холин) происходит в проксимальном сегменте нефрона и обусловлена функционированием специальных систем транспорта. Калий секре-тируется в конечных частях дистального сегмента и собирательных трубках.
Рассмотрим механизм процесса секреции органических кислот на примере выделения почкой ПАГ. При введении ПАГ в кровь человека ее выделение с мочой зависит от фильтрации в клубочках и секреции клетками канальцев (см. рис. 12.5). Когда секреция ПАГ (РАН) достигает максимального уровня (ТтPАН), она становится постоянной и не зависит от содержания ПАГ в плазме крови. Принцип секреторного процесса при транспорте органических соединений состоит в том, что в мембране клетки прокси-
159
мального канальца, обращенной к интерстициальной жидкости, имеется переносчик А, обладающий высоким сродством к ПАГ. В присутствии ПАГ образуется комплекс А—ПАГ, который обеспечивает перемещение ПАГ через мембрану, и на ее внутренней поверхности ПАГ освобождается в цитоплазму. При этом переносчик снова приобретает способность перемещаться к внешней поверхности мембраны и соединяться с новой молекулой ПАГ. Механизм транспорта состоит в том, что переносчик обменивает ПАГ на а-кетоглутарат на базальной плазматической мембране клетки проксимального канальца. Переносчик обеспечивает поступление ПАГ внутрь клетки. Угнетение дыхания цианидами, разобщение дыхания и окислительного фосфорилирования в присутствии динитрофенола снижают и прекращают секрецию. Уровень секреции зависит от числа переносчиков в мембране. Секреция ПАГ возрастает пропорционально увеличению концентрации ПАГ в крови до тех пор, пока все молекулы переносчика не насыщаются ПАГ. Максимальная скорость транспорта ПАГ достигается в тот момент, когда количество ПАГ, доступное для транспорта, становится равным количеству молекул переносчика А, которые могут образовывать комплекс А—ПАГ.
Поступившая в клетку ПАГ движется по цитоплазме к апикальной мембране и с помощью имеющегося в ней специального механизма выделяется в просвет канальца. Способность клеток почки к секреции органических кислот и оснований носит адаптивный характер. Если в течение нескольких дней часто инъецировать ПАГ (или пенициллин), то интенсивность секреции возрастает. Это обусловлено тем, что в клетках проксимальных канальцев при участии систем белкового синтеза вырабатываются вещества, являющиеся необходимыми компонентами процесса переноса через мембрану органических веществ.
Подобно секреции органических кислот, секреция органических оснований (например, холина) происходит в проксимальном сегменте нефрона и характеризуется ТГ1. Системы секреции органических кислот и оснований функционируют независимо друг от друга, при угнетении секреции органических кислот пробенецидом секреция оснований не нарушается.
Транспорт в нефроне К+ характеризуется тем, что К+ не только подвергается обратному всасыванию, но и секретируется клетками эпителия конечных отделов нефрона и собирательных трубок. При реабсорбции из просвета канальца К+ поступает в эпителиальную клетку, где концентрация К+ во много раз выше, чем в канальцевой жидкости, и К+ диффундирует из клетки через базальную плазматическую мембрану в тканевую интерстициаль-ную жидкость, а затем уносится кровью. При секреции К+ поступает в клетку в обмен на Na+ через эту же мембрану с помощью натрий-калиевого насоса, который удаляет Na+ из клетки; тем самым поддерживается высокая внутриклеточная концентрация К+. При избытке К+ в организме система регуляции стимулирует его секрецию клетками канальцев. Возрастает проницаемость для К+
160
мембраны клетки, обращенной в просвет канальца, появляются «каналы», по которым К+ по градиенту концентрации может выходить из клетки. Скорость секреции К+ зависит от градиента электрохимического потенциала на этой мембране клетки: чем больше электроотрицательность апикальной мембраны, тем выше уровень секреции. При введении в кровь и поступлении в просвет канальца слабо реабсорбируемых анионов, например сульфатов, увеличивается секреция К*. Таким образом, секреция К зависит от его внутриклеточной концентрации, проницаемости для КЛ апикальной мембраны клетки и градиента электрохимического потенциала этой мембраны. При дефиците К+ в организме клетки конечных отделов нефрона и собирательных трубок прекращают секрецию К' и только реабсорбируют его из канальцевой жидкости. В этом случае К* из просвета канальца транспортируется через апикальную плазматическую мембрану внутрь клетки, движется по цитоплазме в сторону основания клетки и через базаль-ную плазматическую мембрану поступает в тканевую жидкость, а затем в кровь. Приведенные данные указывают на высокую пластичность клеток этих отделов канальцев, способных под влиянием регуляторных факторов перестраивать свою деятельность, изменяя направление транспорта К+, осуществляя то его реабсорбцню, то секрецию.
Определение величины канальцевой секреции. Секреторную функцию проксимальных канальцев измеряют с помощью веществ, которые выделяются из организма главным образом посредством канальцевой секреции. В кровь вводят ПАГ (или диодраст) вместе с инулином, который служит для измерения клубочковой фильтра ци и. Величина транспорта (Т) органического вещества (Трлн) пРи секреции (S) его из крови в просвет канальца определяется по разности между количеством этого вещества, выделенным почкой {UpAH'V)* и количеством попавшего в мочу вследствие фильтрации в 1С1„*РрАн)*
Приведенная формула характеризует величину секреции вещества почкой при любом уровне загрузки секреторной системы. В то же время мерой работы секреторного аппарата почки служит его максимальная загрузка.
При условии полного насыщения секреторного аппарата ПЛГ определяется величина максимального канальцевого транспорта ПАГ (ТтрАН), которая является мерой количества функционирующих клеток проксимальных канальцев. У человека Тп>лн составляет 80 мг/мин на 1,73 м2 поверхности тела.
Активная секреция органических анионов в проксимальных мочевых канальцах. Эпителиоциты проксимального мочевого канальца секретируют в жидкость канальца большое количество различных эндогенных и экзогенных анионов. См. Таблицу. Некоторые органические анионы, секретируемые в жидкость проксимальных мочевых канальцев. Многие из этих органических анионов уже содержатся в жидкости проксимальных мочевых канальцев. Они попадают туда в результате их фильтрации из плазмы крови капилляров почечного клубочка в полость капсулы Шумлянского-Боумена. Таким образом, количество органических анионов в проксимальном мочевом канальце и их экскреция зависят как от их фильтрации, так и от их секреции. Некоторые из органических анионов по большей части связаны с белками плазмы крови. Такие органические анионы фильтруются лишь в малой степени. Для них секреция является главным механизмом экскреции. Отсюда, максимальная их концентрация в моче может служить характеристикой секреции этих веществ. Механизмы секреции органических анионов в проксимальных мочевых канальцах представляют собой активный транспорт веществ по направлению противоположный реабсорбции органических веществ из жидкости мочевых канальцев. На базолатеральных участках плазмалемм канальцевых эпителиоцитов расположены активные белки-переносчики для транспорта анионов из интерстициальной жидкости в цитоплазму эпителиоцитов. Эти белки-переносчики осуществляют органиченный по скорости транспорт анионов в цитоплазму эпителиоцитов. Транспорт из цитоплазмы эпителиоцитов через апикальные области плазмалемм в жидкость мочевых канльцев осуществляется путем облегчённой диффузии, а также путем активного трансмембранного транспорта. Такой транспорт может осуществляться посредством различных механизмов. Это может быть первичный (однонаправленный, uniport transport, uniport) активный транспорт или вторичный, сопряженный активный транспорт (conjugate transport). Для секреции в мочевых канальцах типичным является сопряженный противонаправленный транспорт (conjugate counter-transport, antiport), зависящий от натрия. Через плотные соединения эпителиоцитов органические анионы (как и глюкоза) практически не транспортируются. Если концентрация определённых органических анионов в плазме крови слишком высока, они не будут эффективно удаляться почками из крови (один из способов удержания в крови необходимой концентрации назначенного медикамента). Относительная неспецифичность белков-переносчиков является причиной их способности удалять из организма большое количество многих экзогенных химических веществ и, в частности, медикаментов. В такой экскреторной функции почки взаимодействуют с печенью. В печени могие экзогенные (и эндогенные) вещества в процессе метаболизма преобразуются и соединяются либо с глюкуронатом, либо с сульфатом. В результате таких преобразований образуются растворимые в воде вещества, удаляемые из плазмы крови путем секреции в мочевых канальцах. Одним из примеров такого способа экскреции органических анионов является секреция парааминогиппурата, используемого для оценки величины эффективного плазматока в почечных тельцах. Секреция парааминогиппурата обеспечивается парой механизмов сопряженного противонаправленного транспорта, по одному с каждой из областей плазмалеммы эпителиоцита. Через базолатеральную область плазмалеммы транспорт аниона парааминогиппурата в цитоплазму эпителиоцита сопряжен с противонаправленным транспортом аниона, образующегося из дикарбоновой кислоты. Через апикальную область плазмалеммы транспорт аниона парааминогиппурата из цитоплазмы эпителиоцита в жидкость мочевого канальца осуществляется другим белком-переносчиком. На начальном этапе секреции с увеличением концентрации парааминогиппурата пропорционально увеличивается скорость потока его секреции (участок линейной зависимости). Постепенно все молекулы белков-переносчиков насыщаются парааминогиппуратом (Tm) и его транспорт замедляется, а затем скорость потока его секреции остается постоянной, независимой (экспонента) от увеличивающейся концентрации парааминогиппурата. Секреция парааминогиппурата является типичным механизмом экскреции органических анионов. Вместе с тем существует много других механизмов экскреции органических анионов.
Ураты.
Ураты, кислые соли мочевой кислоты и их анионы, являются органическими веществами в регулировании содержания которых в жидкостях организма принимают непосредственное участие почки. Нарушение метаболизма пуринов может приводить к ряду заболеваний (подагра, мочекаменная болезнь, болезни иммунодефицита), сопровождающихся изменением нормальной концентрации мочевой кислоты и её солей в жидкостях организма.
Ураты свободно фильтруются. Почти все они реабсорбируются в начальных отделах проксимальных канальцев. Вместе с тем в последующих отделах проксимальных канальцев ураты активно секретируются. Затем, в прямом проксимальном канальце они снова реабсорбируются. Общая скорость потока канальцевой реабсорбции уратов в норме много больше, чем скорость потока их канальцевой секреции. Отсюда массовая скорость потока экскреции составляет лишь небольшую часть массовой скорости потока фильтрации. Существуют различные механизмы транспорта уратов, но преобладающим механизмом является сопряженный с другим органическим анионом противонаправленный транспорт (conjugate counter-transport, antiport). Немотря на то, что реабсорбция уратов преобладает над их секрецией, при увеличении образования уратов (продукт метаболизма пуринов), доля секреции уратов в проксимальных мочевых канальцах увеличивается, что ведет к увеличению их экскеции. При нарушении функций почек экскреция уратов может уменьшаться по трём причинам: (1) уменьшение фильтрации уратов, обусловленное уменьшением объёмной скорости гемациркуляции в капиллярах почечных клубочков, (2) чрезмерная реабсорбция уратов в мочевых канальцах, (3) уменьшение секреции уратов.
Активная секреция органических катионов в проксимальных мочевых канальцах.
В проксимальных мочевых канальцах существует несколько механизмов транспорта органических катионов. Эти механизмы аналогичны механизмам транспорта органических анионов. Разнообразию транспортируемых эндогенных и экзогенных органических катионов соответствует разнообразие белков-переносчиков. См. Таблицу. Некоторые органические катионы, секретируемые в жидкость проксимальных мочевых канальцев.
На начальном этапе секреции, с увеличением концентрации органических катионов пропорционально увеличивается скорость потока их секреции (участок линейной зависимости). Постепенно все молекулы белков-переносчиков насыщаются органическими катионами (Tm) и их транспорт замедляется, а затем скорость потока их секреции остается постоянной, независимой (экспонента) от увеличивающейся концентрации органических катионов.
Механизмы секреции органических анионов в проксимальных мочевых канальцах представляют собой активный транспорт веществ по направлению противоположный реабсорбции органических веществ из жидкости мочевых канальцев. На базолатеральных участках плазмалемм канальцевых эпителиоцитов расположены активные белки-переносчики для транспорта катионов из интерстициальной жидкости в цитоплазму эпителиоцитов. Транспорт из цитоплазмы эпителиоцитов через апикальные области плазмалемм в жидкость мочевых канальцев осуществляется в основном путем активного трансмембранного транспорта (сопряженный противонаправленный транспорт conjugate counter-transport, антипорт antiport). Примером секреции в мочевых канальцах органических катионов является транспорт креатинина.
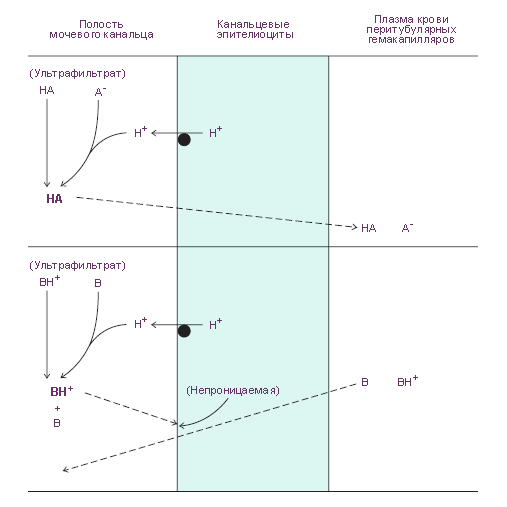
Зависимость пассивной реабсорбции и секреции от рН Многие вещества, содержащиеся в мочевых протоках почек являются слабыми кислотами или слабыми основаниями. Каждое из этих веществ распределено в двух формах: неионизированной (нейтральной) форме и ионизированной форме. Разным значениям рН соответствуют разные доли (в совокупном распределении) неионизированной и ионизированной формы данного вещества. Многие слабые кислоты становятся неионизированными при низких значениях рН . При высоких значениях рН они диссоциируют на анион и протон, то есть переходят в ионизированную форму. Чем меньше рН , тем больше доля неионизированной формы слабой кислоты. И наоборот, Чем больше рН , тем меньше доля неионизированной формы слабой кислоты и больше доля ионизированной (диссоциированной) её формы (анионов).
В общем, липидные плазмалеммы эпителиоцитов мочевых канальцев более проницаемы для неионизированных форм слабых кислот и слабых оснований, чем для их ионизированных форм. Поэтому неионизированные формы этих веществ посредством диффузии могут транспортироваться по концентрационному градиенту через стенку мочевых канальцев в любых направлениях. В то же время для ионизированных форм этих веществ стенка мочевых канальцев является препятствием для диффузии.
Imagine the case in which the
tubular fluid becomes acidified relative to the plasma, which it does on a normal
diet. For a weak acid in the tubular fluid, relatively more will be converted to the
neutral free acid form and, therefore, become more permeable. This favors diffusion
out of the lumen (reabsorption). Therefore, a highly acidic urine (low pH)
tends to increase passive reabsorption of weak acids (and promote less excretion).
For many weak bases, the pH dependence is just the opposite. At low pH they are
protonated cations (trapped in the lumen), whereas at high pH they are converted
to neutral free base. As the urine becomes acidified, more is converted to the impermeable
charged form and is trapped in the lumen. Less is reabsorbed passively,
and more is excreted.
Having said this, what difference does it make? Because so many medically
useful drugs are weak organic acids and bases, all these factors have important
clinical implications. For example, if one wishes to enhance the excretion of a
drug that is a weak acid, one attempts to alkalinize the urine (because this traps
the ionic form in the lumen). In contrast, acidification of the urine is desirable if
one wishes to prevent excretion of the drug. Of course, exactly the opposite applies
to weak organic bases. At any pH, increasing the urine flow increases the excretion
of both weak acids and bases (Figure 5–2). Finally, excretion can be reduced
by administering another drug that interferes with any active proximal secretory
pathway for the drug.
UREA
Urea is a very special substance for the kidney. It is an end product of protein
metabolism, simply waste to be excreted, and also a useful tool for the regulation
of water excretion. Proteins, from which urea is derived, are the action molecules
in cells (eg, transporters and enzymes) and the structural substance of the connective
tissue (eg, collagen). In addition, proteins are the source of metabolic fuel.
Excess dietary protein not needed for tissue synthesis is either oxidized right away
or converted to fat and stored for later oxidation. During fasting, the body breaks
down its own protein for fuel, in essence consuming itself. When oxidized for
fuel, protein is first split into its constituent amino acids. These are then separated
into a nitrogen moiety (ammonium) and a carbohydrate moiety. The carbohydrate
goes on to further metabolic processing, but the ammonium cannot be further
oxidized and is a waste product. The problem is, ammonium is rather toxic to
most tissues (except the medullary interstitium) and the liver immediately converts
it to urea (mostly) and a smaller, but a crucial, amount of glutamine. (We
will take up the fate of this glutamine in Chapter 9 on acid-base balance.) Urea
production proceeds continuously and constitutes about half of the normal solute
content of urine.
The normal level in the blood is quite variable (3 mmol/L–9mmol/L),1 reflecting
variations in both protein intake and renal handling of urea. Over the long
term (days to weeks), renal urea excretion must match hepatic production; otherwise,
plasma levels would rise into the pathological range, producing a condition
1=Plasma urea concentration is usually expressed as blood urea nitrogen (BUN) in units of milligrams
per deciliter. Each molecule of urea contains 2 atoms of nitrogen, so 1 mmol of urea contains 2 mmol of
nitrogen, with a combined weight of 28 mg. Thus, the normal levels of plasma urea are expressed as BUN
values ranging from 8.4 mg/dL to 25.2 mg/dL. We use units of millimoles per liter because we can then
directly convert to osmolality.
called uremia. On a shorter term basis (hours to days), urea excretion rate may
not exactly match production rate because urea excretion is also regulated for
purposes other than keeping a stable plasma level. As discussed in Chapter 6, urea
is a key solute involved in regulating the excretion of water. To summarize renal
handling of urea, urea is freely filtered. About half is reabsorbed in the proximal
tubule. An amount equal to that reabsorbed is then secreted back into the loop of
Henle. Finally, about half is reabsorbed again in the medullary collecting duct.
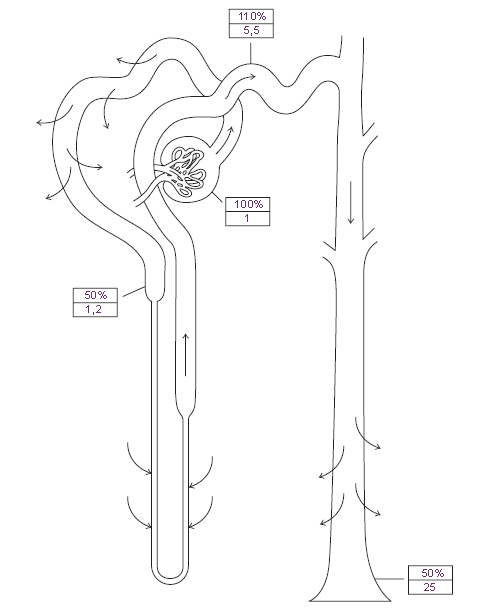
The net result is that about half the filtered load is excreted (Figure 5–3).
As a molecule, urea is small (molecular weight, 60 Da), is water soluble,
and is freely filtered. Because of its highly polar nature, it does not permeate
lipid bilayers, but a set of uniporters (the UT family) transport urea in
various places in the kidney and in other sites within the body (particularly red
blood cells). Because urea is freely filtered, the filtrate contains urea at a concentration
identical to that in plasma. Let us assume a normal plasma level (5 mmol/L).
Roughly half the filtered load is reabsorbed in the proximal tubule. This occurs
primarily by the paracellular route. As water is reabsorbed (about two thirds of the
filtered water is reabsorbed in the proximal tubule), solutes in the lumen that are
not reabsorbed by the transcellular route become concentrated. Urea is prominent
among these solutes. As urea becomes concentrated, it is driven passively through
the leaky tight junctions. By the time the tubular fluid enters the loop of Henle,
about half the filtered urea has been reabsorbed, and the urea concentration has
increased to a little more than its value in the filtrate (because proportionally more
water than urea was reabsorbed). At this point, the process becomes fairly complicated.
First, conditions in the medulla depend highly on the individual’s state of
hydration. Second, there is a difference between superficial nephrons, with short
loops of Henle that only penetrate the outer medulla, and juxtamedullary
nephrons, with long loops of Henle that reach all the way down to the papilla. We
simplify the issue by considering all nephrons together.
The interstitium of the medulla has a considerably higher urea concentration
than plasma. The concentration increases from the outer to the inner medulla
(part of the so-called medullary osmotic gradient), and its peak value in the inner
medulla depends on hydration status and levels of antidiuretic hormone.
We explain these variations in Chapter 6 in connection with regulation of water
excretion. For now, we note that the medullary urea concentration is greater
than in the tubular fluid entering the loop of Henle, so there is a concentration
gradient favoring secretion into the lumen. The tight junctions in the loop
of Henle are no longer permeable, but the epithelial membranes of the thin regions
of the Henle loops express urea uniporters, members of the UT family.
This permits secretion of urea. In fact, the urea secreted from the medullary interstitium
into the thin regions of the loop of Henle replaces the urea previously
reabsorbed in the proximal tubule. Thus, when tubular fluid enters the thick ascending
limb, the amount in the lumen is at least as large as the filtered load.
Because about 80% of the filtered water has now been reabsorbed, the luminal
urea concentration is now several times greater than in the plasma. Beginning
with the thick ascending limb and continuing all the way to the medullary collecting
ducts (through the distal tubule and cortical collecting ducts), the luminal
membrane urea permeability (and the tight junction permeability) is essentially
zero. Therefore, a large amount (roughly the filtered load or more) of urea
is still within the tubular lumen and flowing from the cortical into the medullary
collecting ducts. The concentration is now much greater than in the plasma.
Just how much greater depends on hydration status, because, as is discussed in
Chapter 6, a variable fraction of the remaining water is reabsorbed in the cortical
collecting ducts.
As tubular fluid flows in the collecting-duct system from cortex to medulla,
additional water is reabsorbed. Thus, luminal urea concentration rises even more
and can easily reach 50 times greater than in plasma. We indicated earlier that the
urea concentration in the medullary interstitium is also greater than in plasma,
but the luminal concentration is a little higher, so the gradient favors reabsorption
in the inner medulla. Therefore, urea is reabsorbed for the second time. In fact,
this reabsorbed urea increases medullary interstitial urea concentration and is the
source of urea that is secreted into the loop of Henle. Finally, the result is that half
the original amount of filtered urea passes into the final urine, an amount that,
over the long term, must match hepatic production of urea if the body is to remain
in balance for urea. These processes are summarized in Figure 5–3.
KEY CONCEPTS
The myriad array of organic solutes in the plasma is handled by the kidney; important
metabolites are almost completely reabsorbed (saved), whereas waste products
are, for the most part, excreted.
Most organic solutes are transported transcellularly by a large number of different
saturable multiporters (Tm systems).
Normal filtered loads of glucose are completely reabsorbed by a sodium-glucose
symporter that saturates in conditions of pathological hyperglycemia, leading to
the appearance of glucose in the urine.
Urea is reabsorbed proximally and recycled between the collecting ducts and loop
of Henle in the medulla, resulting in a net excretion of about half the filtered load.
Таблица. Некоторые органические анионы, секретируемые в проксимальных мочевых канальцах. Модификация: Eaton D.C., Pooler J., Vander A.J. Eds. Vander's Renal Physiology. McGraw-Hill Medical, 2002, 215 p. см.: Литература. Иллюстрации.
№
Эндогенные вещества
Экзогенные вещества (медикаменты)
1
Соли жёлчных кислот = Bile salts
Ацетазоламид = Acetazolamide
2
Жирные кислоты = Fatty acids
Хлоротиазид = Chlorothiazide
3
Гиппураты = Hippurates
Этакринат = Ethacrynate
4
Гидроксибензоаты = Hydroxybenzoates
Фуросимид = Furosemide
5
Оксалаты = Oxalate
Пенициллин = Penicillin
6
Простаглдандины = Prostaglandins
Пробенецид = Probenecid
7
Ураты = Urate
Сахарин = Saccharin
8
—
Салицилаты = Salicylates
9
—
Сульфонамиды = Sulfonamides
Таблица. Некоторые органические катионы, секретируемые в проксимальных мочевых канальцах. Модификация: Eaton D.C., Pooler J., Vander A.J. Eds. Vander's Renal Physiology. McGraw-Hill Medical, 2002, 215 p. см.: Литература. Иллюстрации.
№
Эндогенные вещества
Экзогенные вещества (медикаменты)
1
Ацетилхолин = Acetylcholine
Атропин = Atropine
2
Холин = Choline
Изопротеренол = Isoproterenol
3
Креатинин = Creatinine
Циметидин = Cimetidine
4
Дофамин = Dopamine
Меперидин = Meperidine
5
Эпинефрин = Epinephrine
Морфин = Morphine
6
Гуанидин = Guanidine
Прокаин = Procaine
7
Гистамин = Histamine
Хинин = Quinine
8
Serotonin
Тетраэтиламмоний = Tetraethyl ammonium
9
Норэпинефрин = Norepinephrine
—
10
Тиамин = Thiamine
—
Схема. Figure 5–2. Acidification of the luminal fluid creates, by mass action, the gradients that
drive net passive reabsorption of weak acids (top—dashed line) and net passive secretion
of weak bases (bottom—dashed line). The source of the secreted hydrogen ions is
discussed in Chapter 9.. Модификация: Eaton D.C., Pooler J., Vander A.J. Eds. Vander's Renal Physiology. McGraw-Hill Medical, 2002, 215 p. См.: Физиология человека: Литература. Иллюстрации.
Примечание:
Figure 5–2. Acidification of the luminal fluid creates, by mass action, the gradients that
drive net passive reabsorption of weak acids (top—dashed line) and net passive secretion
of weak bases (bottom—dashed line). The source of the secreted hydrogen ions is
discussed in Chapter 9.
Схема. Образование мочи. Модификация: Eaton D.C., Pooler J., Vander A.J. Eds. Vander's Renal Physiology. McGraw-Hill Medical, 2002, 215 p. См.: Физиология человека: Литература. Иллюстрации.
Примечание:
Figure 5–3. Urea handling by the kidney. The arrows indicate the regions where transport
occurs and the direction of transport. The boxes show (in the top half) the percentage
of the filtered load remaining in the tubule and, in the bottom half, the luminal
concentration relative to the concentration in plasma. The numbers are subject to considerable
variability, particularly for regions beyond the proximal tubule, because they
are so dependent on hydration status.
Образование мочи, или мочеобразование - это сложная функция, состоящая из трех взаимодействующихпроцессов, осуществляющихся непосредственно в почках: фильтрация в почечных тельцах нефронов, реабсорбция в канальцах нефронов, секреция в канальцах и собирательных трубочках нефронов. См. схемы 28, 29 из списка иллюстраций на данной странице. Мочеобразование и мочевыведение являются непосредственным или опосредованным результатом ряда других, сопряжённых с функциями почки, функций систем организма.
Впервые концепцию о том, что основой образования мочи является фильтрация веществ из капилляров сосудистого клубочка почечного тельца в полость капсулы почечного тельца сформулировал в 1842 г. германский физиолог К. Людвиг (Карл Фридрих Вильгельм Людвиг, Ludwig Carl Friedrich Wilhelm, 1816-1895, John M. Davis, Klaus Thurau and Dieter Haberle, Carl Ludwig: the discoverer of glomerular filtration, Nephrol Dial Transplant, 1996, 11, 4, 717-720). В первых десятилетиях XX столетия американский фармаколог А.Н. Ричардс (Richards Alfred Newton, 1876-1966) экспериментально подтвердил эту концепцию. С помощью микроманипулятора А.Н. Ричардс микропипеткой проник в полость капсулы почечного тельца и извлёк оттуда ультрафильтрат плазмы крови.
Разрешается некоммерческое цитирование материалов данной энциклопедии при условии полного указания источника заимствования: имени автора, названия и WEB-адреcа данной энциклопедии