|
СЕНСОРНЫЕ РЕЦЕПТОРЫ КОЖИ [ sensory receptors of the skin ] Кожа - это орган тела человека, составляющий поверхность тела, отделяющую тело от его среды и обеспечивающий непосредственное взаимодействие тела со средой.
Кожа предназначена:
(1) для защиты организма от воздействия патогенных факторов среды,
(2) для обмена организма со средой веществами, энергией и информацией,
(3) для поддержания формы тела.
Кожа по массе и по площади поверхности - самый большой орган тела человека. Масса кожи среднего взрослого человека (70 кг, 1,72 м) составляет ~12 ÷ 16% (~4,1 кг) массы тела. Площадь поверхности кожи взрослого человека в среднем составляет ~1,6 м2 (женщины), ~1,8 м2 (мужчины).
В РАЗРАБОТКЕ = UNDER CONSTRUCTION
p. 211
Csillag A., Ed. Atlas of the Sensory Organs: Functional and Clinical Anatomy = Атлас сенсорных органов: функциональная и клиническая анатомия, Humana Press, 2005, 272 p.
Иллюстрированное учебное пособие. В формате .pdf. .
Доступ к данному источнику = Access to the reference.
URL: http://www.tryphonov.ru/tryphonov/serv_r.htm#0 quotation
The Skin and Other Diffuse Sensory Systems
Mihбly Kбlmбn and Andrбs Csillag
ANATOMICAL OVERVIEW OF THE SKIN AND OTHER
DIFFUSE SENSORY SYSTEMS
This chapter summarizes those sensory structures that
have not been covered in Chapters 1 through 4. First, we
provide a general survey of the receptors discussed, then
we describe the skin as a specific organ of sense, followed
by the neural pathways of tactile sensation as exemplified
by the mystacial vibrissae.
Receptors
The classification of receptors is based on the following:
• source of stimulus
• sensation and its biological effect
• morphological structure
• level of perception (conscious or subconscious)
• characteristics of their innervation.
According to their principal structure, the diffuse endings
belong to one of the following three categories:
1. free nerve endings (the major component)
2. encapsulated endings (possessing a lamellar or nonlamellar
connective tissue capsule)
3. epidermal (more generally: cell-associated) endings
Interestingly, the other classifications seem to be independent
of the morphological structure.
According to the source of stimuli, these receptors are
represented in all three main groups: exteroceptors, visceroceptors,
and proprioceptors (the latter two collectively
forming the group of interoceptors).
The exteroceptors (receiving stimulus from the environment)
are represented here by the skin and periorificial
mucosal receptors. For anatomical reasons, some of the
oral mucosal receptors, as well as the corresponding proprioceptors
relevant to the textural attributes of food, have
been described in the chapter on taste sensation.
Cutaneous (skin) receptors are responsible for a multitude
of “skin” sensations. In addition to the classical
“touch,” “warm,” “cold,” and “pain” qualities, we distinguish
hair-bending, pressure, vibration, stretching, itching,
“discomfort,” stroking, tickling, titillation, and
wetness. However, the morphological variety of receptors
seems to be poor when compared to the impressive register
of sensations. The following explanations may be
inferred.
1. the combination of receptor activities (actually, deep
pressure, stretching and vibration cannot be registered
without the participation of the proprioceptive
system, see below);
2. morphologically similar receptors—mainly free
nerve endings—can be used for different stimuli and
sensations;
3. sensation-processing neural systems may modify the
elementary sensations, mainly concerning their pleasant
or unpleasant modalities. The involvement of the
limbic system is to be mentioned here.
Proprioceptors comprise the receptors of the locomotor
system. The term refers to the fact that these receptors are
activated by the active or passive movements of the same
muscles that respond to their stimuli that is, they are
“own” (proprius in Latin) receptors of the muscles. The
vestibular organ may also be classified as a member of
this group, owing to its functional relations. Histologically,
proprioceptors comprise neuromuscular spindles,
neurotendineus organs of Golgi, Pacinian corpuscles, and
bare nerve endings of joints.
Visceroceptors are distributed in the wall of hollow
organs (including blood vessels), the capsule of parenchymal
organs, and the suspension of viscera. They react to
tension and/or muscular contraction rather than touch.
Irritant receptors respond to noxious chemical, mechanical,
or thermal stimuli. Histologically, they correspond to
encapsulated (Pacinian, Krause) or free endings. It has to
be noted, however, that the direct effect of the stimulus
can be similar in the aforementioned systems. Pain can
arise in all of them. Heat can affect the visceral systems,
too, and the mechanical stimuli can always be narrowed
down to compression or stretching. This may explain why
one can find morphologically similar structures in all
three classes of receptors. Functional data also suggest
similar mechanisms of action, e.g., integrin б2в1 may be a
linking agent between mechanical stress in the extracellular
matrix and the modulation of neuronal response. Visceral
chemoreceptors are integrated in the humoral, rather
than the nervous system. Important exceptions are the oxygen
saturation receptors of the aortic and carotid bodies.
According to the electrophysiological characteristics,
the receptive field of receptors may be small or large
(depending on their superficial or deep position), the
threshold may be low or high (the latter is characteristic
for pain), and the adaptation may be slow or rapid (registering
situation or alteration).
The myelinated low threshold cutaneous afferents
belong to four main classes: simple rapidly adapting
(RA), Pacinian corpuscle-associated, rapidly adapting
(PC), slowly adapting, type I (SAI) and SAII.
c. 213=
Иннервация фолликула волоса кожи лица кошки Fig. 5.1 Innervation of hair follicle in the facial skin of cat. Redrawn
after an original silver impregnated specimen of Бbrahбm.
1. Hair shaft
2. Circularly arranged nerve fibers
3. Longitudinally arranged nerve fibers
4. Root sheath
Клетка Меркеля = Fig. 5.2 Schematic drawing of the Merkel cell.
Redrawn and modified after Rцhlich.
1. Keratinocyte
2. Merkel cell
3. Dense core vesicles
4. Lamina basalis
5. Nerve terminal
Тактильное тельце Мейсснера = Fig. 5.3 Schematic drawing of the Meissner’s tactile
corpuscle. Redrawn and modified after Rцhlich.
1. Epithelial
cells
2. Nerve
fibers
3. Myelin
sheaths
Внутренняя капсула тельца Пачини = Fig. 5.4 Schematic drawing of the inner capsule of the Pacinian
corpuscle. Redrawn and modified after Rцhlich.
1. Nerve fiber
2. Concentric cellular
laminae
3. Nucleus of cell
forming the
concentric lamina
Генитальное тельце (аналог тельца Краузе) = Fig. 5.5 Schematic drawing of a genital corpuscle. An essentially
similar organization is found in the end-bulbs of Krause and
various other bulbous “innominate corpuscles.” After Patzelt.
Окончание Гольджи-Маццони = Fig 5.6 Schematic drawing of Golgi-Mazzoni ending. Redrawn and
modified after Rцhlich.
1. Lamellar connective tissue capsule
2. Reticular telodendrion (fine network of axonal
arborization)
3. Nerve fiber
Окончание Руффини (кошка) = Fig 5.7 Reconstructed image of a Ruffini ending in the
hairy skin of cat, composed from serial semithin
sections. Redrawn and modified after Chambers.
Inset: Cross-section of the apical part of Ruffini
ending. Drawing adapted from a serial
electronmicroscopic image of a Ruffini ending in
the hairy skin of cat (Chambers, 1972).
1. Myelinated nerve fiber
2. Endoneural capsule space
3. Endoneurial cell
4. Axon
5. Inner core
6. Collagenous fibrils (see further details on
inset)
7. Capsule
8. Terminal ramification (boutons) of the axon
9. Schwann cell
РЕЦЕПТОРЫ КОЖИ-1. 4 ИЛЛЮСТРАЦИИ
- Схема. Иннервация фолликула волоса кожи лица кошки .
- Схема. Клетка Меркеля.
- Схема. Тактильное тельце Мейсснера.
- Схема. Внутренняя капсула тельца Пачини.
РЕЦЕПТОРЫ КОЖИ-2. 4 ИЛЛЮСТРАЦИИ
- Схема. Генитальное тельце (аналог тельца Краузе).
- Схема. Окончание Гольджи-Маццони.
- Схема. Окончание Руффини (кошка).
- Схема. 2_32 = Gartner Figure 22-1 Various mechanoreceptors. A, Merkel's disk. B, Meissner's corpuscle. C, Pacinian corpuscle. D, Peritricial (naked) nerve endings. E, Ruffini's corpuscle. F, Krause's end bulb. G, Muscle spindle. H, Golgi tendon organ.
Gartner L.P, Hiatt J.M. Color Textbook of Histology, 3th ed., The McGraw-Hill Companies, 2006, 592 p., 446 Ill.
РЕЦЕПТОРЫ КОЖИ-3. 4 ИЛЛЮСТРАЦИИ
- Схема. 2_32 = Gartner Figure 22-2 Pacinian corpuscles (?132). Ca, capsule; IC, inner core; NF, nerve fiber; OC, outer core..
- Схема. 2_32 = Gartner Тактильное тельце Мейсснера = (10_2)Figure 22-3 Meissner's corpuscle (?540). Ca, capsule; N, nuclei; NF, nerve fiber.Клетка Меркеля.
- Схема. 2_93 = Tactile receptors. Skin contains several types of sensory receptors, primarily involved in the sense of touch. In the epidermis are free nerve endings and tactile discs on nerve fibers associated with tactile (Merkel) cells of the basal layer. Both have unencapsulated nerve fibers, as does the root hair plexus around the bases of hair follicles in the dermis. They detect light touch or movement of hair, although epidermal free nerve endings also detect pain and temperature extremes. More complex tactile receptors encapsulated with connective tissue layers are all in the dermis and include tactile (Meissner) receptors (light touch), lamellated (Pacinian) corpuscles (pressure and high-frequency vibration), Krause bulbs (pressure and low-frequency vibrations), and Ruffini corpuscles (continuous pressure and tissue distortion). The latter two receptors are less well characterized structurally and functionally.
= Junqueira L.C., Carneiro J., Eds. Basic Histology: Text and Atlas = Основы гистологии: Текст и атлас, 11th Ed., The McGraw-Hill Companies, 2007. .
- Схема. Внутренняя капсула тельца Пачини.
Камилло Гольджи, Golgi, Camillo, 1843–1926, итальянский врач, цитолог; вместе с испанским гистологом С.Р.Кахалем, Santiago Ramon y Cajal, 1852-1934 в 1906 г. был награжден Нобелевской премией в области физиологии и медицины
Филиппо Пачини, Pacini, Filippo, 1812–1883, итальянский анатом.
Фридрих Зигмунд Меркель, Merkel, Friedrich Sigmund, 1845–1919, германский анатом, гистолог.
Теодор Шванн, Schwann Theodor, 1810–1882, германский физиолог.
Витторио Маццони, Mazzoni, Vittorio, 1880–1940, итальянский физиолог.
Анджело Руффини, Angelo Ruffini, 1864-1929, итальянский гистолог, эмбриолог.
Вильгельм Краузе, Krause Wilhelm, 1833–1910, германский анатом.
Георг Мейснер, Meissner Georg, 1829–1905, германский анатом, гистолог, физиолог.
Абрам Фатер, Vater, Abraham 1684–1751, германский анатом и ботаник.
|
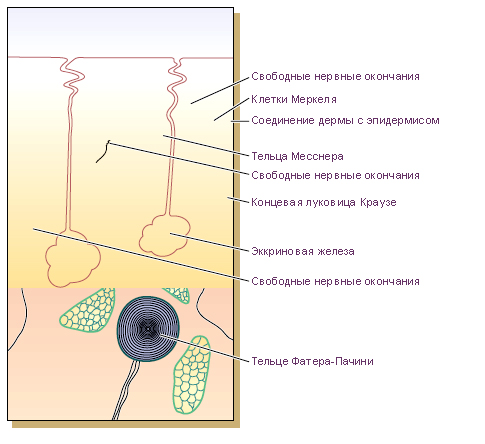
Схема. Нервные окончания в безволосой коже кончика пальца.
Модификация: McGrath J.A., Eady R.A.J., Pope F.M. Anatomy and Organization of Human Skin, 3.1-84. In: Burns D.A., Breathnach S.M., Cox N., Griffiths C.E. Rook's Textbook of Dermatology, 4 vol. set, 7th ed., Wiley-Blackwell, 2004, 4568 p. См.: Физиология человека: Литература. Иллюстрации. |
 |
|
Примечание:
|
Кожа иннервируется приблизительно ~1 миллионом афферентных нервных волокон. Большинство из них заканчивается на лице и верхних конечностях. Наименьшее их количество на спине. Афферентные терминали кожи являются ветвями аксонов клеточных тел, расположенных в спинномозговых (чувствительных) узлах. Диаметр аксонов этих нейронов ~0,2 ÷ 20 мкм. Главный ствол нерва вместе с гемасосудами входит в слой подкожно-жировой клетчатки. Миелинизированные волокна, составляющие нерв, разветвляются в горизонтальную сеть. От неё к поверхности кожи отходят конечные афферентные терминали с многочисленными разветвлениями, переплетающимися в сложную сеть. С уменьшением диаметра нервных волокон уменьшается плотность миелиновой оболочки, образованной Шванновскими клетками. Большинство волокон заканчивается в дерме. И хотя некоторые из нервных волокон пронизывают основную мембрану, они не проходят в эпидермис. Терминальные нервные волокна могут завершаться окончаниями двух типов: (а) сенорными свободными нервными окончаниями и (б) корпускулярными сенсорными не нейрогенными окончаниями. Свободные нервные окончания являются рецепторами, которые не имеют никаких вспомогательных элементов для преобразования воздействий среды в определенную последовательность нервных импульсов, соответствующую воздействию. Корпускулярные сенсорные окончания имеют такие не нейрогенные вспомогательные элементы. Корпускулярные сенсорные окончания подразделяют на инкапсулированные рецепторы и неинкапсулированные рецепторы. Инкапсулированные рецепторы, как правило, располагаются в дерме. Наиболее типичным примером корпускулярных неинкапсулированных рецепторов являются тельца Меркеля. Неинкапсулированные рецепторы, как правило, располагаются в эпидермисе. Наиболее типичным примером инкапсулированных рецепторов являются тельца Пачини.
Тельце Пачини в сечении напоминает луковицу диаметром ~1 мм. Это вспомогательное образование является окончанием миелинизированного сенсорного аксона. Миелиновая оболочка волокна заканчивается в месте входа сенсорного аксона в тельце Пачини.
Заварзин
Фатерпачиииево тельце
В образовании фатерпачиниева тельца принимают участие терминальные разветвления дендрита чувствительной нервной клетки, специализированные фиброциты, система коллагеновых волокон соединительной ткани и видоизмененные глиальные клетки (рис. 145). Наружная капсула образована продольно ориентированными фиброцитами и системой коллагеновых волокон, спирально расположенных вокруг продольной оси. Коллагеновые волокна фиксируются в основании тельца, что обеспечивает лишь незначительные смещения капсулы относительно внутренней части инкапсулированного окончания. Последняя представлена разветвлениями дендритов чувствительных клеток в студенистом аморфном веществе (внутрення колба) и системой глиальных клеток.
В концевых разветвлениях дендритов удается обнаружить большое количество митохондрий. На концевом участке дендрита чувствительной клетки (находящемся во внутренней колбе капсулы) обнаружены синаптические окончания терминалей аксонов. Природа этих аксонов не ясна. Это могут быть окончания отростков вторичночув-ствующих глиальных клеток, которые и воспринимают раздражение своим ресничным аппаратом, как предполагают некоторые морфологи. По мнению физиологов, терминальные разветвления принадлежат отросткам нервных клеток вегетативной нервной системы. Генерация нервного импульса происходит в результате деформации концевого участка дендрита чувствительной клетки. Сложная капсула, образованная фибробластами, коллагеновыми волокнами и глиалькыми клетками, лишь усиливает эффект такой деформации и не принимает непосредственного участия в генерации нервного импульса.
Рис. 145. Строение инкапсулированного интероцептора — тельца Фатер-Пачнни.
/ — слоистая капсула, 2 — внутренняя колба, 3 — дендрит чувствительной нервной клетки спи-
нального ганглия, 4 — спиральные шллагеновые волокна, 5 — фиброциты, 6 — глиальные клетки
с ресничками, 7 — возможные (?) синаптнческне окончания отростков вторичночуаствующих (?)
глиальных клеток или окончания нейронов, регулирующих состояние рецептора.
Тельца Гольджи-Маццони, которые располагаются в гиподерме пальцев человека, таже имеют пластинчатую, но более простую структуру.
Еще одним инкапсулированным рецептором является колбочка Краузе. Колбочки Краузе чаще всего размещаются в поверхностных слоях дермы в местах перехода слизистой в кожу. Колбочки не имеют четко оформленной капсулы. Рыхлый клубочек миелинизированных и немиелинизированных нервных волокон окружен компактной тканью, состоящей из нескольких тонких соединительнотканных пластинок. В этой соединительнотканной оболочке расположена хорошо выраженная сеть гемакапилляров.
Тельца Мейснера длиной ~150 мкм и диаметром ~20 ÷ 40 мкм располагаются на не покрытых волосами участках кожи в сосочках дермы. Эти рецепторы состоят из плоских пластинчатых клеток, сгруппированых в столбик, к которому подходят нервные окончания. Тельце окружено выраженной базальной мембраной и эластичной капсулой.
Тельца Руффини располагаются в суставных капсулах, дерме и гиподерме кожи порытой волосами. Тельце содержит немиелинизированные терминали миелинизированного нервного волокна. Эти терминали оплетают сердцевину тельца, состоящую из коллагеновых волокон. Тельце окружено тонкослойной клеточной капсулой.
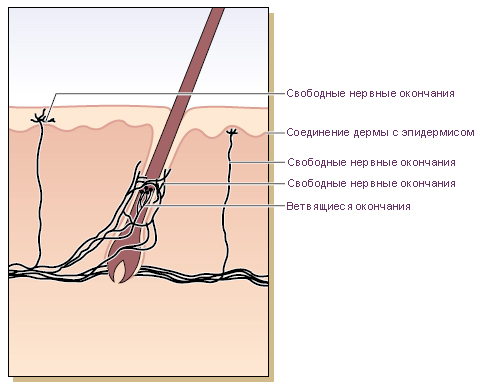
‘Free nerve-endings’, which appear to be derived from
non-myelinated fibres, occur in the superficial dermis and
in the overlying epidermis [9,12,18,19]. Those in the dermis
are arranged in a tuft-like manner and have thus been
designated penicillate nerve endings [18].
Hair follicles have nerve terminals of varying degrees
of complexity (Fig. 3.56). The fine nerve filaments run parparallel
to, and encircle, the hair follicles, forming a palisade.
Each group of axons is surrounded by Schwann cells, and
the side away from the glassy membrane appears to be
free, lacking any specialized contacts with collagenous
fibres [12,21,22]. Sinus hairs, for example the vibrissae of
cats and rats, have a much more complex nerve supply,
with corpuscular as well as non-corpuscular receptor
elements [20,23].
|
|
|
Схема. Нервные окончания в волосистой коже конечностей человека.
Модификация: McGrath J.A., Eady R.A.J., Pope F.M. Anatomy and Organization of Human Skin, 3.1-84. In: Burns D.A., Breathnach S.M., Cox N., Griffiths C.E. Rook's Textbook of Dermatology, 4 vol. set, 7th ed., Wiley-Blackwell, 2004, 4568 p. См.: Физиология человека: Литература. Иллюстрации. |
 |
|
Примечание:
|
Текст общий с предыдущим
|
|
|
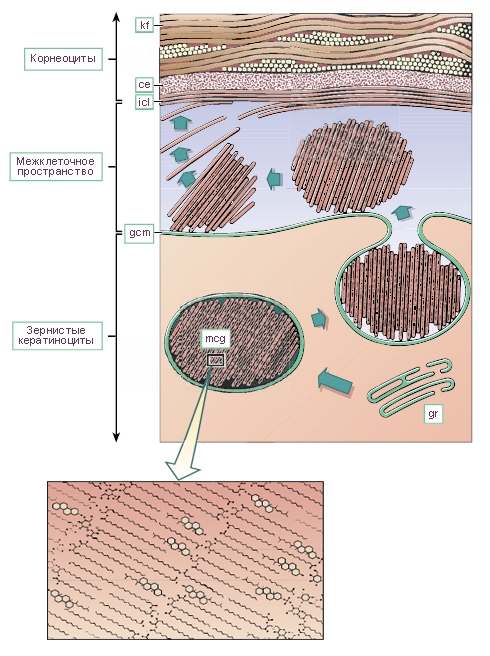
Схема. Образование эпидермального барьера для воды.
Модификация: Archer C.B. Functions of the Skin, 4.1-12. In: Burns D.A., Breathnach S.M., Cox N., Griffiths C.E. Rook's Textbook of Dermatology, 4 vol. set, 7th ed., Wiley-Blackwell, 2004, 4568 p. См.: Физиология человека: Литература. Иллюстрации.
|
 |
Примечание:
Схема создана на основании электронной микрографии. Процесс включает сборку покрытых мембраной гранул - ламеллярных тел (mcg) в области аппарата Гольджи (gr). В последующем гранулы выводятся через плазмалемму (gcm) зернистых кератиноцитов эпидермиса в межклеточное пространство (icl), расположенное между слоем зернистых кератиноцитов и слоем корнеоцитов (клеток рогового слоя). В межклеточном пространстве из гранул высвобождаются кератиновые нити (kf). Они транспортируются в направлении поверхности эпидермиса к внутренней поверхности клеточного конверта (ce) корнеоцита. На вкладке показано увеличенное изображение предполагаемого расположения ацилглюкозилкерамидовых молекул внутри многочисленных бимолекулярных липидных листочков, образованных из липидов гранулярных клеток (кератиноцитов) эпидермиса. В структуре липидов не показаны атомы водорода.
|
|
|
Схема. Пептидная антимикробная защита.
Модификация: Archer C.B. Functions of the Skin, 4.1-12. In: Burns D.A., Breathnach S.M., Cox N., Griffiths C.E. Rook's Textbook of Dermatology, 4 vol. set, 7th ed., Wiley-Blackwell, 2004, 4568 p. См.: Физиология человека: Литература. Иллюстрации. |
 |
|
Примечание:
|
Fig. 4.2 Antimicrobial peptide (AMP) defence. (a) In normal skin,
small amounts of AMPs are produced by the epidermis and are
concentrated around hair follicles and in sweat. (b) After injury or
infection, keratinocytes increase synthesis of AMPs and further
AMPs are produced by neutrophils that are recruited as part of the
acute inflammatory response. (Modified from Gallo et al. [3].) |
|
|
Схема. Расположение сенсорных рецепторов в безволосой и волосистой коже.
Модификация: McGrath J.A., Eady R.A.J., Pope F.M. Anatomy and Organization of Human Skin, 3.1-84. In: Burns D.A., Breathnach S.M., Cox N., Griffiths C.E. Rook's Textbook of Dermatology, 4 vol. set, 7th ed., Wiley-Blackwell, 2004, 4568 p. См.: Физиология человека: Литература. Иллюстрации. |
 |
|
Примечание:
|
Fig. 4.3 Diagram to show the location of sensory receptors in
hairless (glabrous) and hairy skin. The receptors of glabrous skin are
Meissner’s corpuscles and Merkel’s receptors in the dermal papillae,
and bare nerve endings. The receptors of hairy skin are hair-follicle
receptors, Merkel’s receptors, and bare nerve endings. Pacinian
corpuscles and Ruffini’s corpuscles are situated in the subcutaneous
fat and deeper dermis in both glabrous and hairy skin. |
|
См.: Дерматология: Словарь,
Дерматология: Литература. Иллюстрации,
|
«Я У Ч Е Н Ы Й И Л И . . . Н Е Д О У Ч К А ?»
Т Е С Т В А Ш Е Г О И Н Т Е Л Л Е К Т А
Предпосылка:
Эффективность развития любой отрасли знаний определяется степенью соответствия методологии познания - познаваемой сущности.
Реальность:
Живые структуры от биохимического и субклеточного уровня, до целого организма являются вероятностными структурами. Функции вероятностных структур являются вероятностными функциями.
Необходимое условие:
Эффективное исследование вероятностных структур и функций должно основываться на вероятностной методологии (Трифонов Е.В., 1978,..., ..., 2015, …).
Критерий: Степень развития морфологии, физиологии, психологии человека и медицины, объём индивидуальных и социальных знаний в этих областях определяется степенью использования вероятностной методологии.
Актуальные знания: В соответствии с предпосылкой, реальностью, необходимым условием и критерием...
...
о ц е н и т е с а м о с т о я т е л ь н о:
— с т е п е н ь р а з в и т и я с о в р е м е н н о й н а у к и,
— о б ъ е м В а ш и х з н а н и й и
— В а ш и н т е л л е к т !
|
♥ Ошибка? Щелкни здесь и исправь ее! Поиск на сайте E-mail автора (author): tryphonov@yandex.ru
|